

Inwazyjne aspekty biegunek u cieląt. Część II. Biegunki późnego okresu odchowu – kokcydioza
Biologia pasożyta
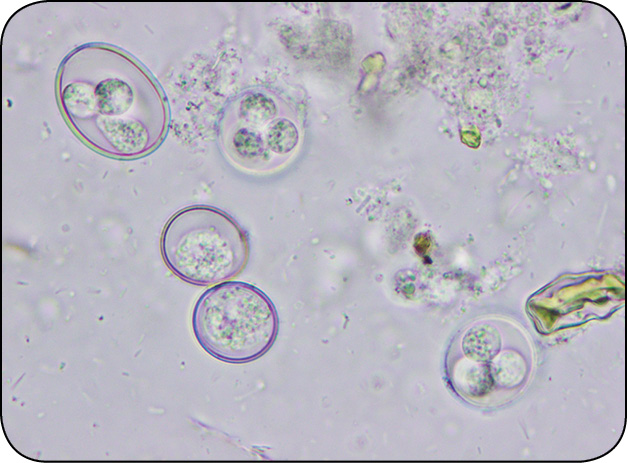
Rodzaj Eimeria klasyfikuje się w grupie tzw. pasożytów monoksenicznych. W cyklu rozwojowym występuje jedynie żywiciel ostateczny, a formy dyspersyjne w postaci oocyst są wydalane do środowiska. W momencie wydalania oocyty są nieinwazyjne. Końcowy etap rozmnażania płciowego pasożyta zachodzi w oocyście (sporogonia) i polega na kolejnych podziałach zygoty, czego efektem jest wytworzenie 4 sporocyst zawierających po 2 sporozoity. Sporozoity są postaciami pasożyta zdolnymi wywołać inwazję, prezentując opisany wyżej bananowaty model budowy pierwotniaka. Oocysty zawierające sporozoity są w pełni inwazyjne. Zarażenie nowego żywiciela następuje na drodze fekalno-oralnej.
Inwazyjne oocyty ze środowiska są zjadane z zanieczyszczonym pokarmem lub wypijane z wodą. Ściana oocyst zostaje nadtrawiona przez enzymy jelita cienkiego, po czym uwolnione do światła jelita sporozoity (dzięki enzymom kompleksu apikalnego) wnikają do komórek nabłonka jelit i tam przechodzą kolejne fazy namnażania. Pierwszą z nich jest faza rozmnażania bezpłciowego na drodze schizogonii. Sporozoity obecne w komórce żywiciela przybierają postać ciała przedpodziałowego (schizonta), w obrębie którego dochodzi do wielokrotnego podziału jądra. Następnie schizont przybiera postać meronta, w którym powstają postacie potomne (merozoity) zawierające jądra po wcześniejszym podziale. Są to też komórki bananowate osiągające 12-16 µm długości.
Gatunki szczególnie patogenne u bydła (E. bovois i E. zuerni) charakteryzują się wytwarzaniem na tym etapie rozwoju szczególnie dużych merontów (makromeronty). Mierzą one do 300 µm średnicy (są wręcz widoczne okiem nieuzbrojonym) i zawierają do 100 000 merozoitów. Gatunki o mniejszej patogenności cechują się na tym etapie rozwoju wytwarzaniem mikromerontów (10 µm średnicy – zawierają do 30 merozoitów). Faza I schizogonii u bydła ma miejsce w komórkach nabłonka jelit cienkich. Na tym etapie nie występują jeszcze istotne objawy kliniczne. W efekcie I schizogonii dochodzi do pękania merontów, a wraz z nimi komórek nabłonka jelit i uwalniania się merozoitów, które kolonizują następne komórki nabłonkowe.
Następuje kolejna, II faza schizogonii. Tym razem liczba komórek objętych inwazją jest wprost proporcjonalna do efektów I schizogonii. II schizogonia E. bovis i E. zuerni ma miejsce w odcinku jelit grubych i w przypadku gatunków patogennych (duża liczba merozoitów I rzędu) wiąże się z uszkodzeniem znacznego odsetka komórek nabłonka. Uwolnione merozoity II generacji wnikają do kolejnych komórek nabłonkowych, przeobrażając się w większości do postaci jednojądrzastych makrogamet. Tylko nieliczne merozoity II generacji po kolonizacji kolejnych enterocytów przechodzą podziały, których skutkiem jest powstanie licznych mikrogamet. Jest to początek etapu rozmnażania płciowego – gamogonii.
Uwolnione z uszkodzonych komórek nabłonkowych mikrogamety (zaopatrzone w trzy wici) wnikają do enterocytów zawierających makrogamety. Następują faza syngamii oraz wytworzenie zygoty. Po rozerwaniu komórki żywiciela zostaje ona otoczona otoczką (powstałą z kwasochłonnych granuli wchodzących w skład makrogamety) i uwalnia się do światła jelita pod postacią oocyty. Na tym etapie kończy się okres prepatentny i rozpoczyna okres patentny inwazji. Jest to o tyle istotne, że od tego momentu jest dopiero możliwa wiarygodna diagnostyka koproskopowa. Rozległe uszkodzenia śluzówki jelit manifestujące się objawami klinicznymi mają miejsce jeszcze w okresie prepatentnym inwazji. Oocysty wydalane z kałem przechodzą dalsze podziały w środowisku zewnętrznym – sporogonia.
Przebieg klinicznych inwazji u bydła jest dość szczególny. Charakteryzują się one relatywnie długim okresem prepatentnym, co jest związane z długotrwałym procesem powstawania makromerontów. II schizogonia u większości gatunków pierwotniaków występujących u innych żywicieli oraz mało patogennych gatunków bydła ma miejsce w 2.-3. dniu po zarażeniu. Proces ten u E. bovis i E. zuerni odbywa się dopiero w 2. tygodniu po zarażeniu, przez co okres prepatentny wydłuża się do 18-21 dni (8, 10, 15, 19, 31, 33).
Patogeneza inwazji
Patogenny wpływ pierwotniaków polega na uszkodzeniu kolonizowanych enterocytów. Mimo że ma ono miejsce już na etapie I schizogonii, to dotyczy ograniczonej liczby komórek. Liczba zarażonych enterocytów wzrasta lawinowo w okresie II schizogonii i jest proporcjonalna do liczby merozoitów I stadium. Gatunki charakteryzujące się obecnością makromerontów w trakcie rozwoju są szczególnie „płodne”, co determinuje zajęcie dużej liczby komórek nabłonka jelit i reakcję w postaci stanu zapalnego. Znacząco uszkadzającym procesem jest także etap gamogonii dotyczący kolejnych partii enterocytów.
Zmiany są obserwowane także w błonie podśluzowej i mięśniowej jelita, które ulegają obrzękom oraz nacieczeniu komórkami związanymi z procesem zapalnym. Generuje to konsekwencje w postaci zaburzenia trawienia oraz wchłaniania wraz z utratą (w przebiegu biegunki): płynów, minerałów, witamin oraz protein. Uszkodzenie naczyń limfatycznych oraz kapilarnych naczyń krwionośnych powoduje dodatkowo krwawienia. U cieląt jest ona szczególnie widoczna, gdyż faza II schizogonii i gamogonii u większości szczególnie patogennych gatunków kokcydiów dotyczy jelit grubych. Dodatkowo uszkodzona błona śluzowa jest bramą wejścia dla innych patogenów (bakterie, wirusy oraz inne pierwotniaki), które wikłają proces chorobowy (2, 5, 7, 9, 10, 12, 13, 17, 33).
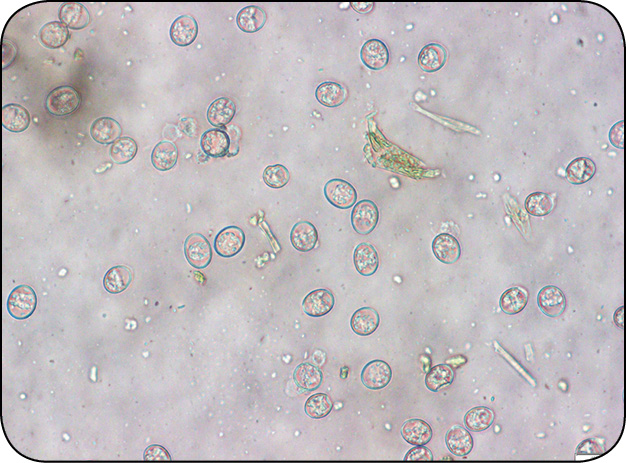
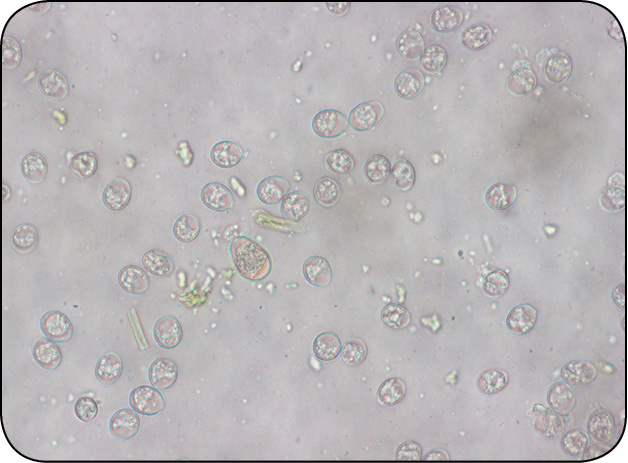
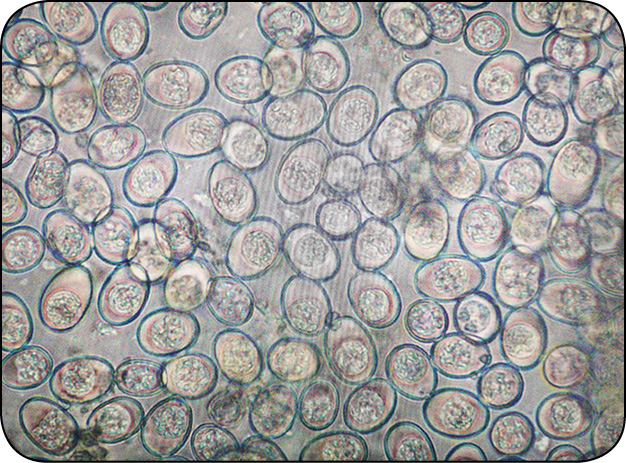
Galeria



POSTĘPOWANIA
w weterynarii





























